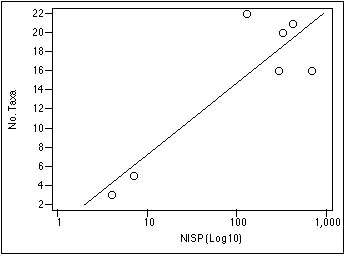
To facilitate the direct comparison of faunal data from excavation units of varying size, I have employed the Volume Control Constant (KV) introduced in Section 3. Multiplying all data from a sample by KV allows me to pretend for analytic purposes that all data derive from uniform excavation units, one square meter in area and 10 cm thick. Such an approach misrepresents the relative taxonomic richness of a sample, which varies as a function of sample size (Grayson 1981:82 - 83). As the number of individual specimens recovered rises, the number of taxa identified should also rise; by manipulating my data with KV , I prevent the direct comparison of taxonomic richness between samples.
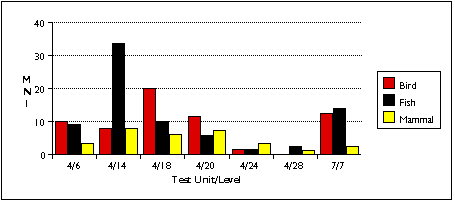
Figure 5.1 demonstrates the relationship of NISP to taxonomic variation. In general, the number of taxa in a unit will rise exponentially as a function of NISP. The moderate correlation between NISP and number of taxa observed (R2 = 0.775) in this instance is probably due to the small number of sample units (n = 7). If I had identified more individuals to finer taxonomic resolution, then the total numbers of taxa identified would also be higher, possibly providing a stronger correlation between NISP and taxonomic richness.
As with the review of analyzed samples in Section 4, unless otherwise noted, all data referred to in Section 5 were derived from analyses of the 1/4" screen sample. Some data from analyses of bulk soil samples were germane to my discussion, however, and I will note the source of these data when I introduce them.
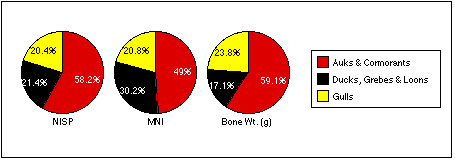
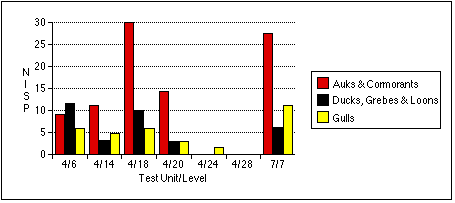
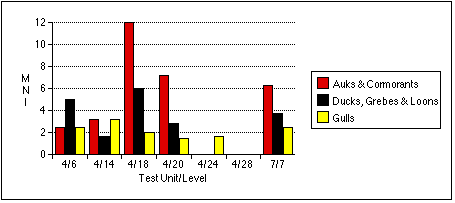
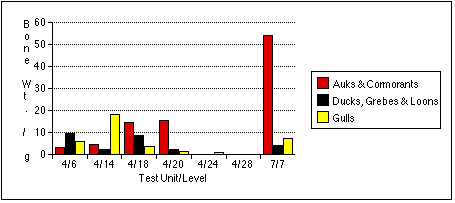
The remains of marsh birds share a similar distribution to cliff birds, although in generally lower numbers. Only sample 4/6 produced the remains of ducks, grebes, and loons in greater relative quantity than those of auks and cormorants by all measures.
Larid remains are the most widely dispersed of all bird types, but often occur in the lowest numbers. Gull remains are the most abundant by NISP in sample 7/7, but more abundant by MNI and bone weight in sample 4/14. Larid bones occur in roughly equal frequencies throughout the other samples; they are the only birds identified in the Stratum IV soils from Test Unit 4.
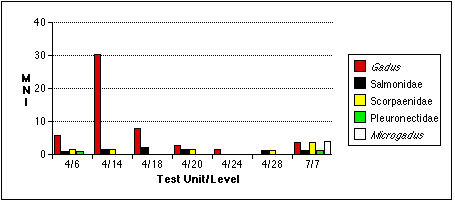
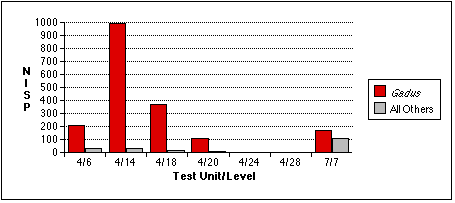
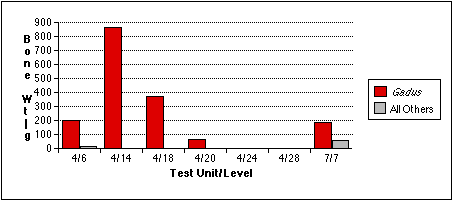
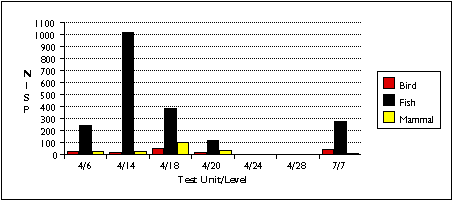
Gadus bones also tend to occur in distinct concentrations (Figure 5.5a - c) . They are especially concentrated in sample 4/14, an unsurprising result given the discretionary nature of my sample selection process. I had targeted this unit precisely because it contained a preponderance of fish bone of an apparently uniform variety, and because the excavation level intersected with the fish bone midden deposit, Stratum II-9. Cod remains also predominate in samples 4/6, 4/14, and 4/18. Gadus remains are less dominant in the TU7/7 sample, where they are the most abundant fish by bone weight and NISP, but second to Pacific tomcod in MNI. The relative abundance of Gadus and other fish remains in TU7/7 may be skewed by the small sample size of this sample; if the fish bone sample recovered from TU7/7 were larger, non-Gadus fish might appear even more abundant. Other fish taxa eclipse Gadus in TU4, Level 28. Cod were completely absent from this sample, which contained the remains of a single salmonid and a single scorpaenid.
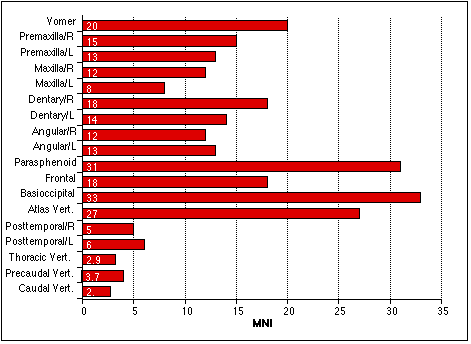
Three elements appear to vary from the skewed distribution of cranial and postcranial elements: left and right posttemporals and the atlas vertebra. In gadids, pectoral girdle bones such as the posttemporal actually lie posterior to the base of the head (as indicated by the articular surface of the basioccipital; Cannon 1987:45), so I consider these bones marginal to the cranium. When I compute postcranial MNI using left posttemporals (potentially the most abundant postcranial element), this lowers the cranial to postcranial MNI ratio to 33:6, or 5.5. The anomalous distribution of atlas vertebrae is best explained by noting that atlases are the first vertebrae immediate posterior to the basioccipital; each Gadus has only one atlas vertebrae (Cannon 1987). If the distribution of cranial and postcranial elements in the analyzed samples is related to a butchering practice involving the beheading of cod, then I would expect that the atlas would represent the most abundant vertebral element, and a better indicator of cranial MNI than postcranial MNI.
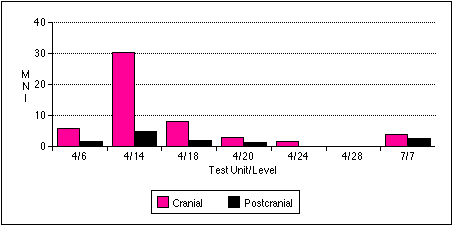
I did not observe the differential distribution of Gadus macrocephalus heads and bodies equally through all sample units, however (Figure 5.7) . When adjusted for excavation volume, sample 4/14 has the highest ratio of cranial to postcranial elements: 30.4 individuals computed using the most abundant cranial element versus 4.8 computed using vertebral elements, or 6.3 times more cod heads than cod bodies. Conversely, sample 7/7 has nearly equal distributions of cranial and postcranial elements: at least 3.75 crania and 2.5 vertebral columns, or only 1.5 times more cod heads than cod bodies.
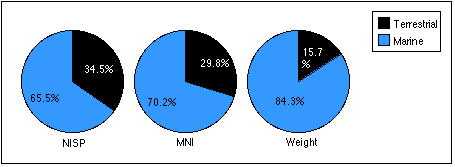
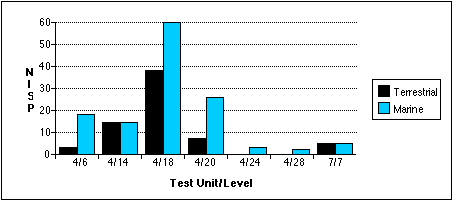
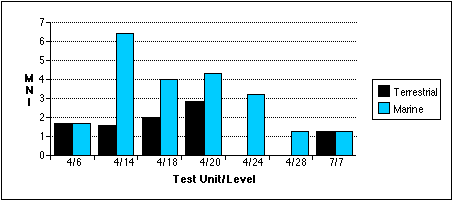
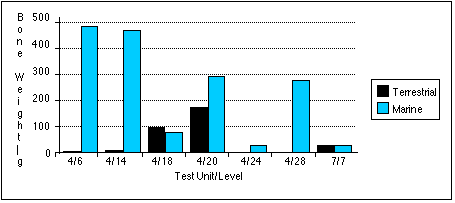
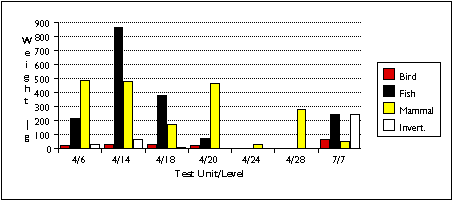
With the exception of the TU7, Level 7 sample, marine mammal remains outnumber terrestrial remains in all units by most measures (Figures 5.9 a - c) . Land mammal remains outweigh marine mammal remains in the TU4, Level 18 sample, probably because much of the marine mammal material recovered from this level belonged to a small juvenile phocid.
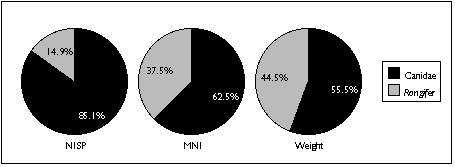
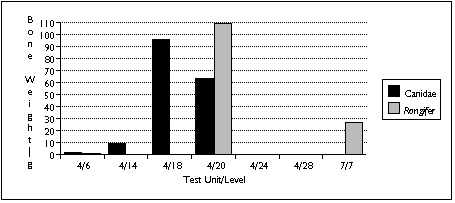
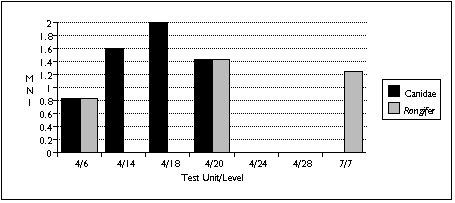
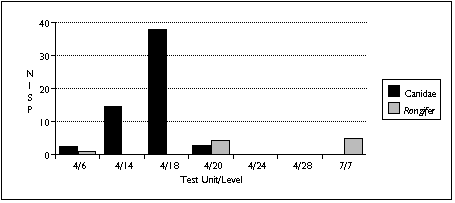
The difficulty of distinguishing dog species archaeologically complicates interpretation of the canid remains. Four varieties of canid are known to be found on Nunivak Island: domestic dog, wolf, and two species of fox (Lantis 1946). I distinguished between these species in my sample mainly by size. Wolves probably contributed the largest identified canid specimens, found in the TU4/20 sample; foxes probably contributed the smallest--several teeth found in TU4/6. I assumed that other canid remains probably belonged to the domestic dog.
Similary, Rangifer remains could either belong to caribou or reindeer. Caribou populations fluctuated widely on Nunivak Island, and mainland Yupiit armed with high-power rifles exterminated these animals in the late 19th century. Reindeer were introduced by American traders early in the 20th century to replace the extinct caribou (Griffin 1996). The distinction between these two species therefore serves as an important temporal marker--but without Rangifer in the comparative collection, I was unable to distinguish between them. Based on radiocarbon dates and a lack of associated historic artifacts, however, I assumed that all Rangifer remains predate the 20th century, and belong to caribou.
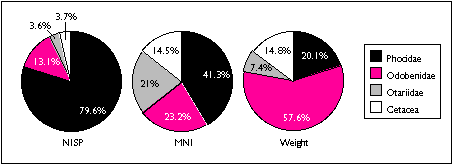
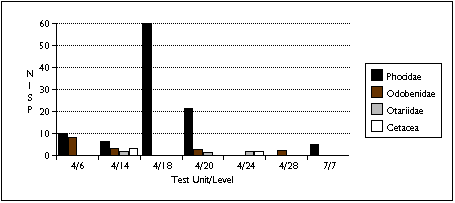
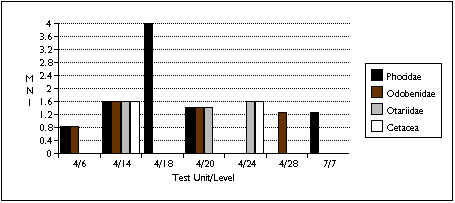
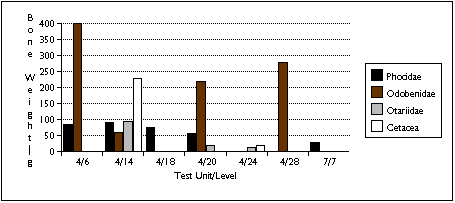
Limited sample size and area of excavation may explain the lack of strong patterning in the distribution of marine mammal remains (Figure 5.13 a - c) . Phocid remains are the most widely distributed, found in five units. Phocids were also the most concentrated: after adjusting for excavated volume, TU4/18 contained 60 phocid specimens and four individuals. Odobenid remains were also spread throughout the sample, with remains found in four sample units. However, I characterize the sea mammal sample as patchily distributed, and add the caveat that this may reflect sample size rather than a particular Ellikarrmiut hunting or processing pattern.
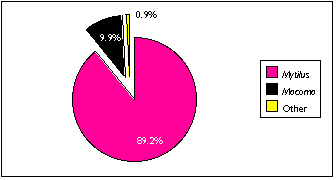
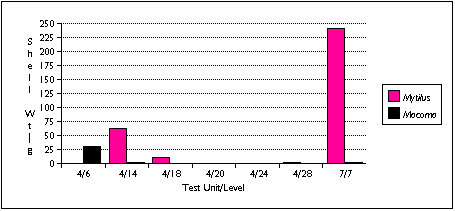
Virtually all shellfish remains were found in the TU7/7 sample; this certainly reflected a bias in sample selection as well as the actual distribution of shellfish (Figure 5.15) . TU7 contained the largest concentration of shell midden material discovered by archaeologists at Nash Harbor, and I included one sample from TU7 to provide some data on invertebrate harvesting patterns. I also noted a large amount of Mytilus shell in the fish bone midden deposit intersecting TU4/14, and I identified several Macoma shell fragments in the TU4/6 sample.
The analysis of invertebrates almost certainly under-represents their actual abundance at Nash Harbor. Screeners noted amounts of Mytilus periostraca during excavation; many of these remains were crushed during screening, or blown out of the screens by the strong Nunivak winds. The use of 1/4" screens during excavation also led to the loss of an unknown amount of shell, which was often so finely fragmented that it passed through the excavation screens.
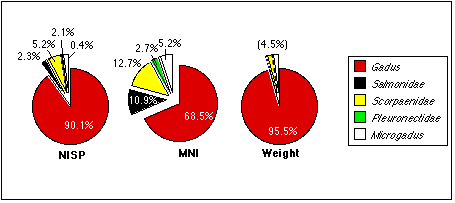
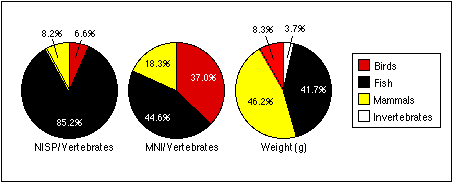
Variations in the size of the different taxa partly explain these contradictory data. Except for Mustelidae, the smallest mammal individual would outweigh the largest individual of any other taxon. The remains of a single walrus individual, represented only by a few fragments, outweighs dozens of more complete bird skeletons. Bird bones, on the other hand, were less fragmented than mammal bones, and easier to identify generally. While I could identify 68.5% of the bird specimens, I could only identify 55.1% of the mammal specimens. Thus, the MNI for birds would necessarily be higher than for other taxa.
After careful consideration of the spatial distribution of faunal remains, I can characterize the analyzed samples with the following summary statements:
No taxon predominated in TU4/6 , except that Gadus --as in other units where fish were found in significant quantities--were clearly the most abundant fish. Except for a few Macoma , invertebrate remains were generally scarce. The mammalian sample was typified by a few large Odobenus specimens.
TU4/14 represented a trash heap of fish bones, associated with a processing activity resulting in the differential deposition of Gadus crania here. Remains of other taxa, especially gulls, cliff birds, and sea mammals, were also abundant.
TU4/18 represented a deposit of fish and bird bones. As with TU4/14, Gadus crania were more numerous than vertebrae. Cliff birds' remains were most concentrated in this sample. Seal and canid predominated among mammalian remains, although a few fragments of whale bone made Cetacea more abundant by bone weight. The presence of Clupea harengus vertebrae in the bulk sample from TU4/18 suggests that herring may also be abundant in this unit, but I can not estimate their relative abundance.
Faunal materials were more scarce in TU4/20 than in the analyzed samples above it. A few Odobenus specimens outweighed all other animal remains, but Phocidae were the most common sea mammals by NISP. Cliff birds remained the most common avians.
Faunal materials were almost entirely lacking in TU4/24 and TU4/28 , except for a few large Odobenus specimens in TU4/28. Notably, these units contained no Gadus remains. The paucity of trash in these levels may bolster the hypothesis that this stratum represented a collapsed structure, as advanced in Section 2.
Mytilus remains predominated in TU7/7 , almost exceeding fish remains in weight. Gadus remains were common, but unlike other samples containing Gadus , cranial and postcranial individuals were nearly equally represented. Quite a few cliff birds were found here as well. I characterize TU7/7 as associated with shellfish and cliff bird processing.
Section 4: Review of Analyzed Sample
Section 6: Ellikarrmiut Economy